- Joined
- Nov 19, 2002
- Messages
- 2,107
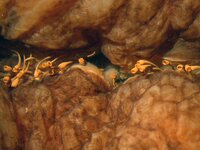
Hey guys, sorry for floating this thread and then apparently abandoning it. I am still puzzling this out, and have been corresponding with a few other ceph-heads about it. The latest development is that Dick Young was in Bergen, where he examined the Type of Chaunoteuthis mollis, and took the attached photo for me of... the same kind of implanted spermatophores! (Actually, spermatangia - and thanks for posting that animation, that was very cool.) C. mollis appears to be a spent Onychoteuthis, but we don't know what species yet - tentacles are lost of course, so the beaks need examining.
I like the theory about the sperm being released through (or creating) the external bulb-like structures, especially since the implanted spermatangia in the male O. meridiopacifica don't have this structure, and wouldn't have released the sperm.
I like the theory about the sperm being released through (or creating) the external bulb-like structures, especially since the implanted spermatangia in the male O. meridiopacifica don't have this structure, and wouldn't have released the sperm.